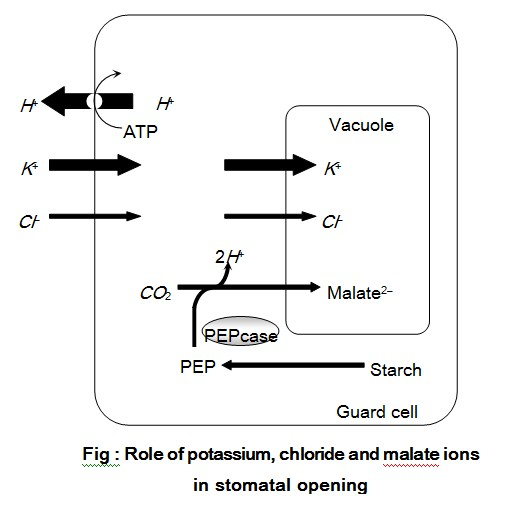
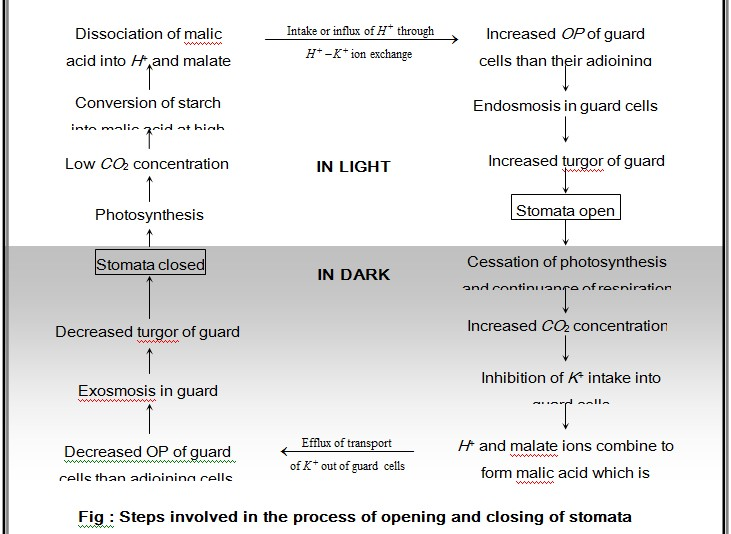
(i) External factors
# (a) Atmospheric humidity : If the atmosphere is humid, it reduces the rate of transpiration. When the air is dry, the rate of transpiration increases.
# (b) Temperature : It affects the rate of transpiration only indirectly. Increase in the temperature of the air decreases the humidity of the air and therefore more water is vapourised and lost from the transpiring surface. The lowering of the air-temperature, on the other hand, increases the humidity and rate of water-loss as well.
# (c) Light : Light affects the rate of transpiration due to its effect on temperature and photosynthesis. During daytime stomata open wide but during night they close. Moreover, during the daytime the light also helps in raising the temperature. Thus increased temperature and presence of wide open stomata increase the rate of transpiration. Light is the most important factor in the regulation of transpiration.
# (d) Atmospheric pressure : The rate of transpiration is inversely proportional to the atmospheric pressure.
# (e) Available soil water : If the available water in the soil is not sufficient the rate of transpiration is decreased. Under internal water deficiency the stomata are partially or completely closed.
# (f) Wind velocity : A transpiring surface of leaf continuously adds water vapours to the atmospheric air. Once the immediate area becomes saturated, it reduces the rate of transpiration. Wind velocity removes the air of that area, which is replaced by fresh air and result in an increases in the rate of transpiration. Wind velocity is measured by anemometer.
(ii) Internal factors/Plant factors
# (a) Leaf area : If leaf area is more, transpiration is faster. However, the rate of transpiration per unit area is more in smaller leaves than in larger leaves due to high number of stomata in a small leaf. Number of stomata per unit area of leaf is called stomatal frequency.
here, I = Stomatal index
S = No. of stomata per unit area
E = No. of epidermal cells in unit area.
# (b) Leaf structure : The anatomical features of leaves like sunken or vestigial stomata; presence of hair, cuticle or waxy layer on the epidermis; presence of hydrophilic substances such as gums, mucilage etc. in the cells; compactly arranged mesophyll cells etc. help in reducing the rate of transpiration.
# (c) Root shoot ratio : According to Parker (1949) the rate of transpiration is directly proportional to the root-shoot ratio.
# (d) Age of plants : Germinating seeds show a slow rate of transpiration. It becomes maximum at maturity. However, it decreases at senescence stage.
# (e) Orientation of leaves : If the leaves are arranged transversely on the shoot they lose more water because they are exposed to direct sunlight. If placed perpendicularly they transpire at slower rate.
Significance of transpiration : The advantages and disadvantages of transpiration are discussed below :
# (i) Advantages
(a) Transpiration is important for plants because it directly influences the absorption of water from the soil.
(b) Transpiration exerts a tension or pull on water column in xylem which is responsible for the ascent of sap.
(c) Transpiration helps in the movement of water and minerals absorbed by the roots to the other parts of the plant.
(d) The evaporation of water during transpiration contributes to the cooling of leaves (and also the surrounding air) and protects leaves from heat injury particularly under conditions of high temperature and intense sunlight.
# (ii) Disadvantages
(a) Transpiration often results in water deficit which causes injury to the plants by desiccation.
(b) Rapid transpiration causes mid-day leaf water deficit (temporary wilting). If such condition continues for some time, permanent water deficit (permanent wilting) may develop, which causes injury to plants.
(c) Many xerophytes have to develop structural modifications to reduce transpiration. These modifications are extra burden on the plants.
(d) Excessive rate of transpiration leads to stunted growth of plants.
(e) Deciduous trees have to shed their leaves during autumn to check transpiration.
(f) Since approximately 90 percent of absorbed water is lost through transpiration, the energy used in absorption and conduction of water goes waste.
Besides all the above mentioned disadvantages, the process of transpiration is unavoidable, because of the anatomical structure of the leaves. Since stomata are required for gaseous exchange in photosynthesis and respiration, the loss of water through them cannot be avoided. Therefore, Curtis (1926) truely called 'transpiration as a necessary evil'.
Anti-transpirants : Most of the water absorbed by plants is lost to the atmosphere by transpiration and hence water use by plants is very inefficient. In recent years efforts have been made to improve the efficiency of water use by the plants. One of the approaches is to reduce transpiration by the application of certain chemical substances. 'The chemical substances which reduce transpiration (by increasing leaf resistance to water vapour diffusion) without affecting gaseous exchange, are called anti-transpirants'. Anti-transpirants are of two types metabolic inhibitors and film forming anti-transpirants.
#(i) Metabolic inhibitors : They reduce transpiration by causing partial closure of stomata, without influencing other metabolic processes, the most important of these inhibitors are phenyl mercuric acetate (PMA) and abscissic acid (ABA).
# (ii) Film forming anti-transpirants : They check transpiration by forming a thin transparent film on the transpiring surface. They are sufficiently permeable to carbon dioxide and oxygen to allow photosynthesis and respiration, but prevent movement of water vapour through them. The important chemicals of this group are silicon emulsion, colourless plastic resins and low viscosity waxes.
(i) External factors
# (a) Atmospheric humidity : If the atmosphere is humid, it reduces the rate of transpiration. When the air is dry, the rate of transpiration increases.
# (b) Temperature : It affects the rate of transpiration only indirectly. Increase in the temperature of the air decreases the humidity of the air and therefore more water is vapourised and lost from the transpiring surface. The lowering of the air-temperature, on the other hand, increases the humidity and rate of water-loss as well.
# (c) Light : Light affects the rate of transpiration due to its effect on temperature and photosynthesis. During daytime stomata open wide but during night they close. Moreover, during the daytime the light also helps in raising the temperature. Thus increased temperature and presence of wide open stomata increase the rate of transpiration. Light is the most important factor in the regulation of transpiration.
# (d) Atmospheric pressure : The rate of transpiration is inversely proportional to the atmospheric pressure.
# (e) Available soil water : If the available water in the soil is not sufficient the rate of transpiration is decreased. Under internal water deficiency the stomata are partially or completely closed.
# (f) Wind velocity : A transpiring surface of leaf continuously adds water vapours to the atmospheric air. Once the immediate area becomes saturated, it reduces the rate of transpiration. Wind velocity removes the air of that area, which is replaced by fresh air and result in an increases in the rate of transpiration. Wind velocity is measured by anemometer.
(ii) Internal factors/Plant factors
# (a) Leaf area : If leaf area is more, transpiration is faster. However, the rate of transpiration per unit area is more in smaller leaves than in larger leaves due to high number of stomata in a small leaf. Number of stomata per unit area of leaf is called stomatal frequency.
here, I = Stomatal index
S = No. of stomata per unit area
E = No. of epidermal cells in unit area.
# (b) Leaf structure : The anatomical features of leaves like sunken or vestigial stomata; presence of hair, cuticle or waxy layer on the epidermis; presence of hydrophilic substances such as gums, mucilage etc. in the cells; compactly arranged mesophyll cells etc. help in reducing the rate of transpiration.
# (c) Root shoot ratio : According to Parker (1949) the rate of transpiration is directly proportional to the root-shoot ratio.
# (d) Age of plants : Germinating seeds show a slow rate of transpiration. It becomes maximum at maturity. However, it decreases at senescence stage.
# (e) Orientation of leaves : If the leaves are arranged transversely on the shoot they lose more water because they are exposed to direct sunlight. If placed perpendicularly they transpire at slower rate.
Significance of transpiration : The advantages and disadvantages of transpiration are discussed below :
# (i) Advantages
(a) Transpiration is important for plants because it directly influences the absorption of water from the soil.
(b) Transpiration exerts a tension or pull on water column in xylem which is responsible for the ascent of sap.
(c) Transpiration helps in the movement of water and minerals absorbed by the roots to the other parts of the plant.
(d) The evaporation of water during transpiration contributes to the cooling of leaves (and also the surrounding air) and protects leaves from heat injury particularly under conditions of high temperature and intense sunlight.
# (ii) Disadvantages
(a) Transpiration often results in water deficit which causes injury to the plants by desiccation.
(b) Rapid transpiration causes mid-day leaf water deficit (temporary wilting). If such condition continues for some time, permanent water deficit (permanent wilting) may develop, which causes injury to plants.
(c) Many xerophytes have to develop structural modifications to reduce transpiration. These modifications are extra burden on the plants.
(d) Excessive rate of transpiration leads to stunted growth of plants.
(e) Deciduous trees have to shed their leaves during autumn to check transpiration.
(f) Since approximately 90 percent of absorbed water is lost through transpiration, the energy used in absorption and conduction of water goes waste.
Besides all the above mentioned disadvantages, the process of transpiration is unavoidable, because of the anatomical structure of the leaves. Since stomata are required for gaseous exchange in photosynthesis and respiration, the loss of water through them cannot be avoided. Therefore, Curtis (1926) truely called 'transpiration as a necessary evil'.
Anti-transpirants : Most of the water absorbed by plants is lost to the atmosphere by transpiration and hence water use by plants is very inefficient. In recent years efforts have been made to improve the efficiency of water use by the plants. One of the approaches is to reduce transpiration by the application of certain chemical substances. 'The chemical substances which reduce transpiration (by increasing leaf resistance to water vapour diffusion) without affecting gaseous exchange, are called anti-transpirants'. Anti-transpirants are of two types metabolic inhibitors and film forming anti-transpirants.
#(i) Metabolic inhibitors : They reduce transpiration by causing partial closure of stomata, without influencing other metabolic processes, the most important of these inhibitors are phenyl mercuric acetate (PMA) and abscissic acid (ABA).
# (ii) Film forming anti-transpirants : They check transpiration by forming a thin transparent film on the transpiring surface. They are sufficiently permeable to carbon dioxide and oxygen to allow photosynthesis and respiration, but prevent movement of water vapour through them. The important chemicals of this group are silicon emulsion, colourless plastic resins and low viscosity waxes.