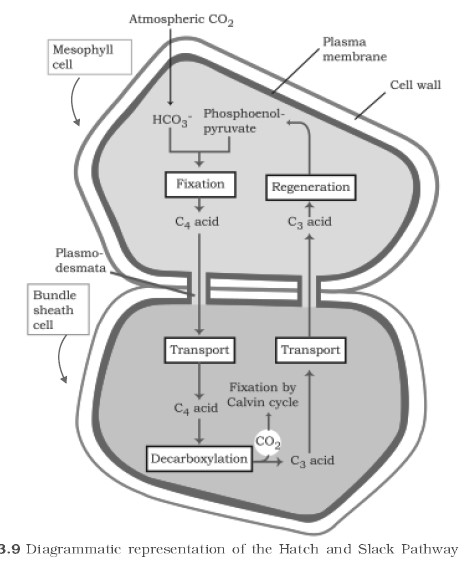
Kortschak and Hart supplied CO2 to the leaves of sugarcane, they found that the first stable product is a four carbon (C4) compound oxalo acetic acid instead of 3-carbon atom compound. The detailed study of this cycle has introduced by M.D. Hatch and C.R. Slack (1966). So it is called as "Hatch and Slack cycle". The stable product in C4 plant is dicarboxylic group. Hence it is called dicarboxylic acid cycle or DCA-cycle. C4 plants are true xerophytic plants.
The plants that perform C4 cycle are found in tropical (Dry and hot regions) and sub-tropical regions. There are more than 900 known species in which C4 cycle occurs. Among them, more than 300 species belong to dicots and the rest belong to monocots. The important among them are sugarcane, maize, Sorghum, Cyperus rotundus, Digitaria brownii, Amaranthus, etc. These plants have "Kranz" (German term meaning halo or wreath) type of leaf anatomy. The vascular bundles, in C4 leaves are surrounded by a layer of bundle sheath cells that contain large number of chloroplasts. The chloroplasts in C4 leaves are dimorphic (Two morphologically distinct types). The chloroplasts of bundle sheath cells are larger in size and arranged centripetally. They contain starch grains but lack grana. The mesophyll cells, on the other hand, contain normal types of chloroplasts. The mesophyll cells perform C4 cycle and the cells of bundle sheath perform C3 cycle.
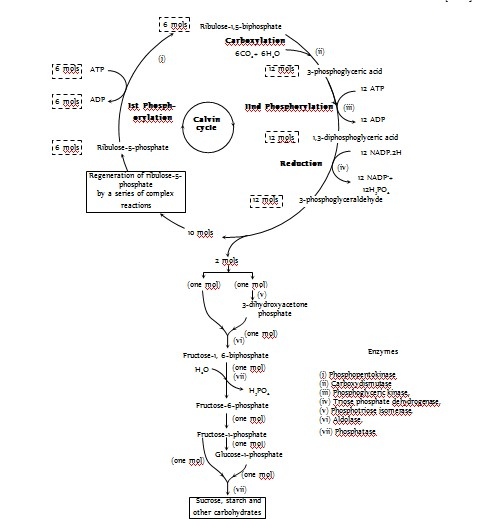
CO2 taken from the atmosphere is accepted by phosphoenolpyruvic acid (PEP) present in the chloroplasts of mesophyll cells of these leaves, leading to the formation of a 4-C compound, oxaloacetic acid (OAA). This acid is converted to another 4-C acid, the malic acid which enters into the chloroplasts of bundle sheath cells and there undergoes oxidative decarboxylation yielding pyruvic acid (a 3-C compound) and CO2. CO2 released in bundle sheath cells reacts with Ribulose-1,5-biphosphate (RuBP) already present in the chloroplasts of bundle sheath cells and thus Calvin cycle starts from here. Pyruvic acid re-enters mesophyll cells and regenerates phosphoenol pyruvic acid. CO2 after reacting with RuBP gives rise to sugars and other carbohydrates. Mesophyll cells have PEP carboxylase and pyruvate orthophosphate dikinase enzyme while the bundle sheath cells have decarboxylase and complete enzymes of Calvin cycle. In C4 plants, there are 2 carboxylation reactions, first in mesophyll chloroplast and second in bundle sheath chloroplast.
C4 plants are better photosynthesizers. There is no photorespiration in these plants. In C4 plants, for formation of one mole of hexose (glucose) 30 ATP and 12 NADPH2 are required. There is difference in different C4 plants in mechenism of C4 mode of photosynthesis. The main difference is in the way the 4C dicarboxylic acid is decarboxylated in the bundle sheath cells.
# The three categories of C4 pathways in C4 plants are recognised such as :
(a) Some C4 plants e.g., Zea mays, Saccharum officinarum, Sorghum utilise NADP+ specific malic enzyme for decarboxylation. This mechanism of C4 pathway in these C4 plants is said to be of NADP+ –Me Type.
(b) Some C4 plants e.g., Atriplex, Portulaca, Amaranthus utilise NAD+ specific malic enzyme for decarboxylation. This mechanism of C4 pathways in these C4 plants is said to be of NAD+ –Me Type.
(c) Some C4 plants e.g., Panicum, Chloris utilise PEP-carboxykinase enzyme. The mechanism of C4 pathway in these plants is called as PCK-me-Type.
# Characteristics of C4 cycle
(1) C4 species have greater rate of CO2 assimilation than C3 species. This is on account of the fact that
(a) PEP carboxylase has great affinity for CO2.
(b) C4 plants show little photorespiration as compared to C3 plants, resulting in higher production of dry matter.
(2) C4 plants are more adapted to environmental stresses than C3 plants.
(3) CO2 fixation by C4 plants require more ATP than that by C3 plants. This additional ATP is needed for conversion of pyruvic acid to phosphoenol pyruvic acid and its transport.
(4) CO2 acceptor molecule in C4 plants is PEP. Further, PEP-carboxylase (PEPCO) is the key enzyme (RuBP-carboxylase enzyme is negligible or absent in mesophyll chloroplast, but is present in bundle sheath chloroplast).
Kortschak and Hart supplied CO2 to the leaves of sugarcane, they found that the first stable product is a four carbon (C4) compound oxalo acetic acid instead of 3-carbon atom compound. The detailed study of this cycle has introduced by M.D. Hatch and C.R. Slack (1966). So it is called as "Hatch and Slack cycle". The stable product in C4 plant is dicarboxylic group. Hence it is called dicarboxylic acid cycle or DCA-cycle. C4 plants are true xerophytic plants.
The plants that perform C4 cycle are found in tropical (Dry and hot regions) and sub-tropical regions. There are more than 900 known species in which C4 cycle occurs. Among them, more than 300 species belong to dicots and the rest belong to monocots. The important among them are sugarcane, maize, Sorghum, Cyperus rotundus, Digitaria brownii, Amaranthus, etc. These plants have "Kranz" (German term meaning halo or wreath) type of leaf anatomy. The vascular bundles, in C4 leaves are surrounded by a layer of bundle sheath cells that contain large number of chloroplasts. The chloroplasts in C4 leaves are dimorphic (Two morphologically distinct types). The chloroplasts of bundle sheath cells are larger in size and arranged centripetally. They contain starch grains but lack grana. The mesophyll cells, on the other hand, contain normal types of chloroplasts. The mesophyll cells perform C4 cycle and the cells of bundle sheath perform C3 cycle.
CO2 taken from the atmosphere is accepted by phosphoenolpyruvic acid (PEP) present in the chloroplasts of mesophyll cells of these leaves, leading to the formation of a 4-C compound, oxaloacetic acid (OAA). This acid is converted to another 4-C acid, the malic acid which enters into the chloroplasts of bundle sheath cells and there undergoes oxidative decarboxylation yielding pyruvic acid (a 3-C compound) and CO2. CO2 released in bundle sheath cells reacts with Ribulose-1,5-biphosphate (RuBP) already present in the chloroplasts of bundle sheath cells and thus Calvin cycle starts from here. Pyruvic acid re-enters mesophyll cells and regenerates phosphoenol pyruvic acid. CO2 after reacting with RuBP gives rise to sugars and other carbohydrates. Mesophyll cells have PEP carboxylase and pyruvate orthophosphate dikinase enzyme while the bundle sheath cells have decarboxylase and complete enzymes of Calvin cycle. In C4 plants, there are 2 carboxylation reactions, first in mesophyll chloroplast and second in bundle sheath chloroplast.
C4 plants are better photosynthesizers. There is no photorespiration in these plants. In C4 plants, for formation of one mole of hexose (glucose) 30 ATP and 12 NADPH2 are required. There is difference in different C4 plants in mechenism of C4 mode of photosynthesis. The main difference is in the way the 4C dicarboxylic acid is decarboxylated in the bundle sheath cells.
# The three categories of C4 pathways in C4 plants are recognised such as :
(a) Some C4 plants e.g., Zea mays, Saccharum officinarum, Sorghum utilise NADP+ specific malic enzyme for decarboxylation. This mechanism of C4 pathway in these C4 plants is said to be of NADP+ –Me Type.
(b) Some C4 plants e.g., Atriplex, Portulaca, Amaranthus utilise NAD+ specific malic enzyme for decarboxylation. This mechanism of C4 pathways in these C4 plants is said to be of NAD+ –Me Type.
(c) Some C4 plants e.g., Panicum, Chloris utilise PEP-carboxykinase enzyme. The mechanism of C4 pathway in these plants is called as PCK-me-Type.
# Characteristics of C4 cycle
(1) C4 species have greater rate of CO2 assimilation than C3 species. This is on account of the fact that
(a) PEP carboxylase has great affinity for CO2.
(b) C4 plants show little photorespiration as compared to C3 plants, resulting in higher production of dry matter.
(2) C4 plants are more adapted to environmental stresses than C3 plants.
(3) CO2 fixation by C4 plants require more ATP than that by C3 plants. This additional ATP is needed for conversion of pyruvic acid to phosphoenol pyruvic acid and its transport.
(4) CO2 acceptor molecule in C4 plants is PEP. Further, PEP-carboxylase (PEPCO) is the key enzyme (RuBP-carboxylase enzyme is negligible or absent in mesophyll chloroplast, but is present in bundle sheath chloroplast).