Energy Relations - ATP Generation
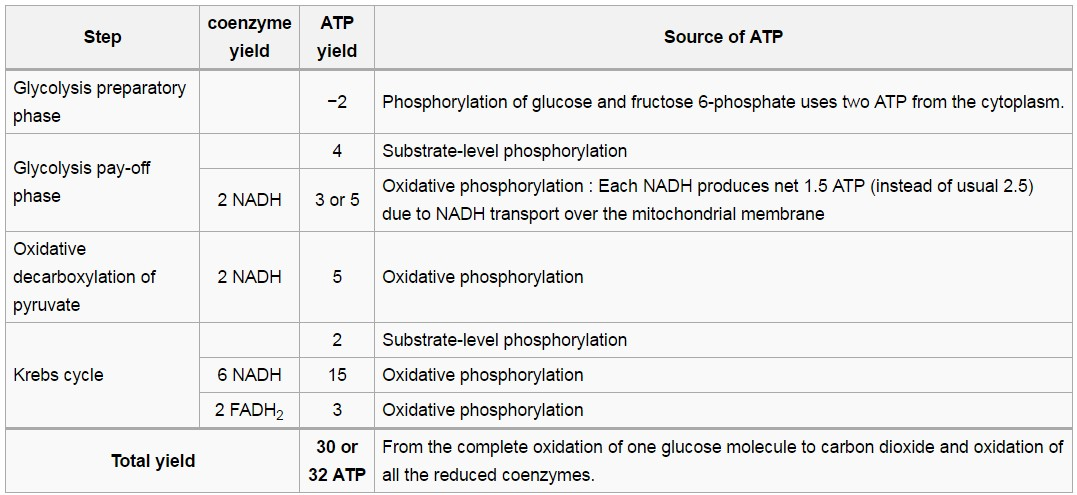
The table below describes the reactions involved when one glucose molecule is fully oxidized into carbon dioxide. It is assumed that all the reduced coenzymes are oxidized by the electron transport chain and used for oxidative phosphorylation.
Although there is a theoretical yield of 38 ATP molecules per glucose during cellular respiration, such conditions are generally not realized because of losses such as the cost of moving pyruvate (from glycolysis), phosphate, and ADP (substrates for ATP synthesis) into the mitochondria. All are actively transported using carriers that utilize the stored energy in the proton electrochemical gradient.
# Pyruvate is taken up by a specific, low Km transporter to bring it into the mitochondrial matrix for oxidation by the pyruvate dehydrogenase complex.
# The phosphate carrier (PiC) mediates the electroneutral exchange (antiport) of phosphate (H2PO4−; Pi) for OH− or symport of phosphate and protons (H+) across the inner membrane, and the driving force for moving phosphate ions into the mitochondria is the proton motive force.
# The ATP-ADP translocase (also called adenine nucleotide translocase, ANT) is an antiporter and exchanges ADP and ATP across the inner membrane. The driving force is due to the ATP (−4) having a more negative charge than the ADP (−3), and thus it dissipates some of the electrical component of the proton electrochemical gradient.
The outcome of these transport processes using the proton electrochemical gradient is that more than 3 H+ are needed to make 1 ATP. Obviously this reduces the theoretical efficiency of the whole process and the likely maximum is closer to 28–30 ATP molecules. In practice the efficiency may be even lower because the inner membrane of the mitochondria is slightly leaky to protons. Other factors may also dissipate the proton gradient creating an apparently leaky mitochondria. An uncoupling protein known as thermogenin is expressed in some cell types and is a channel that can transport protons. When this protein is active in the inner membrane it short circuits the coupling between the electron transport chain and ATP synthesis. The potential energy from the proton gradient is not used to make ATP but generates heat. This is particularly important in brown fat thermogenesis of newborn and hibernating mammals.
Stoichiometry of aerobic respiration and most known fermentation types in eucaryotic cell. Numbers in circles indicate counts of carbon atoms in molecules, C6 is glucose C6H12O6, C1 carbon dioxide CO2. Mitochondrial outer membrane is omitted.
According to some of newer sources the ATP yield during aerobic respiration is not 36–38, but only about 30–32 ATP molecules / 1 molecule of glucose, because:
# ATP : NADH+H+ and ATP : FADH2 ratios during the oxidative phosphorylation appear to be not 3 and 2, but 2.5 and 1.5 respectively. Unlike in the substrate-level phosphorylation, the stoichiometry here is difficult to establish.
# ATP synthase produces 1 ATP / 3 H+. However the exchange of matrix ATP for cytosolic ADP and Pi (antiport with OH− or symport with H+) mediated by ATP–ADP translocase and phosphate carrier consumes 1 H+ / 1 ATP as a result of regeneration of the transmembrane potential changed during this transfer, so the net ratio is 1 ATP : 4 H+.
# The mitochondrial electron transport chain proton pump transfers across the inner membrane 10 H+ / 1 NADH+H+ (4 + 2 + 4) or 6 H+ / 1 FADH2 (2 + 4).
So the final stoichiometry is
1 NADH+H+ : 10 H+ : 10/4 ATP = 1 NADH+H+ : 2.5 ATP
1 FADH2 : 6 H+ : 6/4 ATP = 1 FADH2 : 1.5 ATP
# ATP : NADH+H+ coming from glycolysis ratio during the oxidative phosphorylation is :
- 1.5, as for FADH2, if hydrogen atoms (2H++2e−) are transferred from cytosolic NADH+H+ to mitochondrial FAD by the glycerol phosphate shuttle located in the inner mitochondrial membrane.
- 2.5 in case of malate-aspartate shuttle transferring hydrogen atoms from cytosolic NADH+H+ to mitochondrial NAD+
So finally we have, per molecule of glucose.
# Substrate-level phosphorylation: 2 ATP from glycolysis + 2 ATP (directly GTP) from Krebs cycle
# Oxidative phosphorylation
-> 2 NADH+H+ from glycolysis: 2 × 1.5 ATP (if glycerol phosphate shuttle transfers hydrogen atoms) or 2 × 2.5 ATP (malate-aspartate shuttle)
-> 2 NADH+H+ from the oxidative decarboxylation of pyruvate and 6 from Krebs cycle: 8 × 2.5 ATP
-> 2 FADH2 from the Krebs cycle: 2 × 1.5 ATP
Altogether this gives 4 + 3 (or 5) + 20 + 3 = 30 (or 32) ATP per molecule of glucose
The total ATP yield in ethanol or lactic acid fermentation is only 2 molecules coming from glycolysis, because pyruvate is not transferred to the mitochondrion and finally oxidized to the carbon dioxide (CO2), but reduced to ethanol or lactic acid in the cytoplasm.
Although there is a theoretical yield of 38 ATP molecules per glucose during cellular respiration, such conditions are generally not realized because of losses such as the cost of moving pyruvate (from glycolysis), phosphate, and ADP (substrates for ATP synthesis) into the mitochondria. All are actively transported using carriers that utilize the stored energy in the proton electrochemical gradient.
# Pyruvate is taken up by a specific, low Km transporter to bring it into the mitochondrial matrix for oxidation by the pyruvate dehydrogenase complex.
# The phosphate carrier (PiC) mediates the electroneutral exchange (antiport) of phosphate (H2PO4−; Pi) for OH− or symport of phosphate and protons (H+) across the inner membrane, and the driving force for moving phosphate ions into the mitochondria is the proton motive force.
# The ATP-ADP translocase (also called adenine nucleotide translocase, ANT) is an antiporter and exchanges ADP and ATP across the inner membrane. The driving force is due to the ATP (−4) having a more negative charge than the ADP (−3), and thus it dissipates some of the electrical component of the proton electrochemical gradient.
The outcome of these transport processes using the proton electrochemical gradient is that more than 3 H+ are needed to make 1 ATP. Obviously this reduces the theoretical efficiency of the whole process and the likely maximum is closer to 28–30 ATP molecules. In practice the efficiency may be even lower because the inner membrane of the mitochondria is slightly leaky to protons. Other factors may also dissipate the proton gradient creating an apparently leaky mitochondria. An uncoupling protein known as thermogenin is expressed in some cell types and is a channel that can transport protons. When this protein is active in the inner membrane it short circuits the coupling between the electron transport chain and ATP synthesis. The potential energy from the proton gradient is not used to make ATP but generates heat. This is particularly important in brown fat thermogenesis of newborn and hibernating mammals.
Stoichiometry of aerobic respiration and most known fermentation types in eucaryotic cell. Numbers in circles indicate counts of carbon atoms in molecules, C6 is glucose C6H12O6, C1 carbon dioxide CO2. Mitochondrial outer membrane is omitted.
According to some of newer sources the ATP yield during aerobic respiration is not 36–38, but only about 30–32 ATP molecules / 1 molecule of glucose, because:
# ATP : NADH+H+ and ATP : FADH2 ratios during the oxidative phosphorylation appear to be not 3 and 2, but 2.5 and 1.5 respectively. Unlike in the substrate-level phosphorylation, the stoichiometry here is difficult to establish.
# ATP synthase produces 1 ATP / 3 H+. However the exchange of matrix ATP for cytosolic ADP and Pi (antiport with OH− or symport with H+) mediated by ATP–ADP translocase and phosphate carrier consumes 1 H+ / 1 ATP as a result of regeneration of the transmembrane potential changed during this transfer, so the net ratio is 1 ATP : 4 H+.
# The mitochondrial electron transport chain proton pump transfers across the inner membrane 10 H+ / 1 NADH+H+ (4 + 2 + 4) or 6 H+ / 1 FADH2 (2 + 4).
So the final stoichiometry is
1 NADH+H+ : 10 H+ : 10/4 ATP = 1 NADH+H+ : 2.5 ATP
1 FADH2 : 6 H+ : 6/4 ATP = 1 FADH2 : 1.5 ATP
# ATP : NADH+H+ coming from glycolysis ratio during the oxidative phosphorylation is :
- 1.5, as for FADH2, if hydrogen atoms (2H++2e−) are transferred from cytosolic NADH+H+ to mitochondrial FAD by the glycerol phosphate shuttle located in the inner mitochondrial membrane.
- 2.5 in case of malate-aspartate shuttle transferring hydrogen atoms from cytosolic NADH+H+ to mitochondrial NAD+
So finally we have, per molecule of glucose.
# Substrate-level phosphorylation: 2 ATP from glycolysis + 2 ATP (directly GTP) from Krebs cycle
# Oxidative phosphorylation
-> 2 NADH+H+ from glycolysis: 2 × 1.5 ATP (if glycerol phosphate shuttle transfers hydrogen atoms) or 2 × 2.5 ATP (malate-aspartate shuttle)
-> 2 NADH+H+ from the oxidative decarboxylation of pyruvate and 6 from Krebs cycle: 8 × 2.5 ATP
-> 2 FADH2 from the Krebs cycle: 2 × 1.5 ATP
Altogether this gives 4 + 3 (or 5) + 20 + 3 = 30 (or 32) ATP per molecule of glucose
The total ATP yield in ethanol or lactic acid fermentation is only 2 molecules coming from glycolysis, because pyruvate is not transferred to the mitochondrion and finally oxidized to the carbon dioxide (CO2), but reduced to ethanol or lactic acid in the cytoplasm.