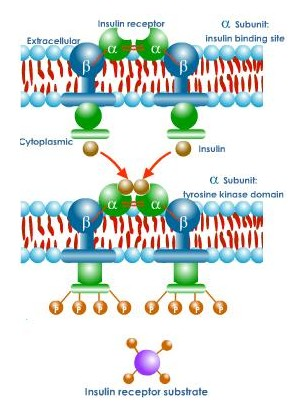
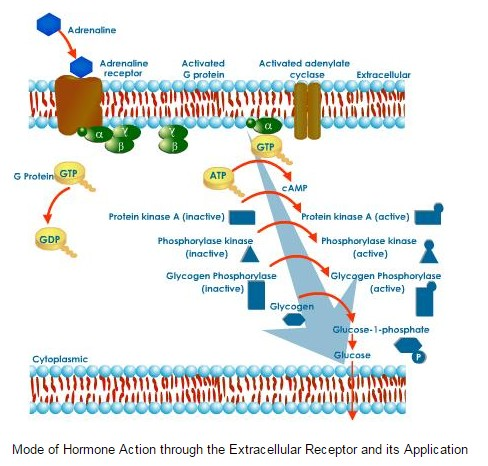
Mediators amplify the signal in an expanding cascade of response. A single -subunit of insulin receptor, for example, activates many molecules of DG, and each protein kinase C molecule activated by DG will, in turn, activate many other enzyme molecules. DG and IP3 are examples of second messengers, intermediary compounds that amplify a hormonal signal and so set into action a variety of events within the affected cell. A variety of events within the affected cell. A variety of hormones use another second messenger, the cyclic form of adenosine monophosphate, (cAMP). The enzyme adenylate cyclase converts adenosine triphosphate (ATP) into cAMP. Because an enzyme can be used over and over again, a single molecule of active adenylate cyclase can catalyse production of about 100 molecules of cAMP. In muscle or liver cells, when hormones, such as, adrenaline bind receptors, the receptors change shape and bind to G protein, causing it, in turn, to bind the nucleotide guanosine triphosphate (GTP) and activate another protein adenylate cyclase. The result of this complex cascade of interactions is the production of large amounts of cAMP.
cAMP activates the enzyme protein kinase A, which, in turn, activates the enzyme phosphorylate kinase. Each molecule of protein kinase A activates roughly 100 molecules of enzyme, phosphorylate kinase and so on. The net result is that a single molecule of adrenaline may lead to release of as many as 100 million molecules of glucose within only 1 or 2 minutes. No wonder only very small quantities of hormone are needed.
Mediators amplify the signal in an expanding cascade of response. A single -subunit of insulin receptor, for example, activates many molecules of DG, and each protein kinase C molecule activated by DG will, in turn, activate many other enzyme molecules. DG and IP3 are examples of second messengers, intermediary compounds that amplify a hormonal signal and so set into action a variety of events within the affected cell. A variety of events within the affected cell. A variety of hormones use another second messenger, the cyclic form of adenosine monophosphate, (cAMP). The enzyme adenylate cyclase converts adenosine triphosphate (ATP) into cAMP. Because an enzyme can be used over and over again, a single molecule of active adenylate cyclase can catalyse production of about 100 molecules of cAMP. In muscle or liver cells, when hormones, such as, adrenaline bind receptors, the receptors change shape and bind to G protein, causing it, in turn, to bind the nucleotide guanosine triphosphate (GTP) and activate another protein adenylate cyclase. The result of this complex cascade of interactions is the production of large amounts of cAMP.
cAMP activates the enzyme protein kinase A, which, in turn, activates the enzyme phosphorylate kinase. Each molecule of protein kinase A activates roughly 100 molecules of enzyme, phosphorylate kinase and so on. The net result is that a single molecule of adrenaline may lead to release of as many as 100 million molecules of glucose within only 1 or 2 minutes. No wonder only very small quantities of hormone are needed.